看啥推荐读物

专栏名称: 生信宝典
| 生物信息分析入门、晋级和经验分享。Linux、R、Python学习教程;高通量测序数据分析学习教程;生信软件安装教程。所有内容均为原创分享,致力于从基础学习到提高整个过程。 |
今天看啥
微信公众号rss订阅, 微信rss, 稳定的RSS源
目录
相关文章推荐

|
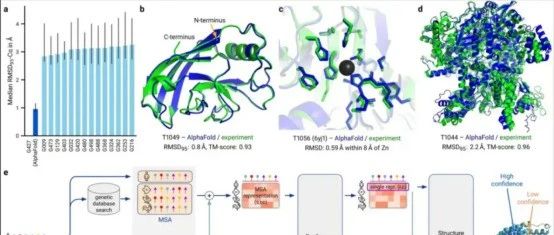
BioArt · Cell Rep | ...· 3 天前 |
|
生信人 · 史上最全格局打开,细讲基因组学,确定不来学学?· 3 天前 |

|
BioArt · 深圳湾实验室申雪桐-沈卫军课题组联合博士后招聘· 4 天前 |

|
生物探索 · Science | ...· 5 天前 |

|
生信宝典 · 5 篇 NAR | ...· 6 天前 |

推荐文章
|
|
生信人 · 史上最全格局打开,细讲基因组学,确定不来学学? 3 天前 |
|
|
BioArt · 深圳湾实验室申雪桐-沈卫军课题组联合博士后招聘 4 天前 |

|
CROU制药在线 · GMP管理体系的搭建改善及变更、偏差实施策略解析 1 年前 |

|
多知 · 张一鸣向家乡福建省龙岩市捐赠5亿元成立“芳梅教育发展基金” 2 年前 |

|
就那点小破事儿 · 最高效的工作关系是每周只见14小时? 4 年前 |
